発表論文から
ペチュニアにおける花弁の模様はRNAサイレンシングによって形成される
Koseki, M., Goto, K., Masuta, C. and Kanazawa, A. (2005) The star-type color pattern inPetunia hybrida‘Red Star’ flowers is induced by the sequence-specific degradation of the chalcone synthase RNA.Plant Cell Physiol.46, 1879-1883.

RNA干渉が医療に使える可能性がでてきたことも関連して、RNAサイレンシングに関する研究は2000年前後からいっそう盛んになりました。RNAサイレンシングの機構の一つとして、転写後にRNAが塩基配列特異的な分解を受けることは、1990年にカルコーン合成酵素
(CHS) 遺伝子を導入したペチュニアを用いた研究によって明らかになりました。その後、さまざまな生物で、外来遺伝子や二本鎖RNAを導入することで同様な現象がおきることが示されました。では、外来の核酸を導入していない細胞では、こうしたRNAサイレンシングはおきうるのでしょうか?
ペチュニアのレッドスターという品種は、その名のとおり、赤色地に白色の星型の模様の花を咲かせます。園芸植物であるペチュニアの育種は1830年代に欧州の人が南米から持ち帰った野生の植物を交配したことに始まります。レッドスターのような美しい模様を持った植物は、ペチュニアの育種の歴史の中で、その初期から作られていましたが、なぜ、そのような模様が作られるのかは解明されていませんでした。私たちの研究グループでは、この論文で、レッドスターの星型模様の形成が、アントシアニン色素を合成するCHS遺伝子の転写後のRNA分解によっておきていることを証明しました。遺伝子特異的な転写後のRNA分解によって内在性遺伝子の発現が制御されていることを証明した例は、microRNAによる制御を除いては、このレッドスターの例を含めて、まだ数例しかありません。
この論文は、Faculty of 1000において、以下のように紹介されました。
“This paper confirms the long-held suspicion that the red and white patterned
petals of the commercial petunia variety 'Red Star' result from an RNAi-type
mechanism that specifically degrades the mRNA of the pigment gene chalcone
synthase (CHS) in unpigmented sectors. Thus, prior to the discovery of cosuppression
(or RNAi) ofCHSby sense transgenes in petunia, plant breeders selected a naturally occurring
case of this important gene silencing mechanism.” (Marjori Matzke, Austrian
Academy of Sciences, Austria)
レッドスターにおけるRNAサイレンシングについては、次世代シーケンサーを利用した研究(下記)により、外来遺伝子によって引き起こされるRNAサイレンシング(コサプレッション)とRNA分解によって生じるたくさんの低分子RNAに共通性があること、すなわち、RNA分解の様式自体に共通性があることを明らかにしました。
Kasai, M., Matsumura, H., Yoshida, K., Terauchi, R., Taneda, A. and Kanazawa,
A. (2013) Deep sequencing uncovers commonality in small RNA profiles between
transgene-induced and naturally occurring RNA silencing of chalcone synthase-A
gene in petunia.BMC Genomics14, 63.
なお、私たちの研究を含む、ペチュニアを用いたRNAサイレンシングに関する研究の歴史については、Dr. Ortrun Mittelsten Scheidが以下の総説において解説しています。
Mittelsten Scheid I (2019) Illuminating (white and) purple patches.Plant Cell31: 128-1209.
真核生物がどのようにして、このようなRNAサイレンシングの機構を獲得したのかは、たいへん興味深い問題です。 RNAサイレンシングは、細胞の中で遺伝子発現の交通整理を行うポリスマンなのでしょうか!?

環境に対する適応に寄与したダイズのレトロトランスポゾン
Liu, B., Kanazawa, A., Matsumura, H., Takahashi, R., Harada, K. and Abe,
J. (2008) Genetic redundancy in soybean photoresponses associated with
duplication of the phytochrome A gene. Genetics 180, 995-1007.
Kanazawa, A., Liu, B., Kong, F., Arase, S. and Abe, J. (2009) Adaptive
evolution involving gene duplication and insertion of a novel Ty1/copia-like retrotransposon in soybean. J. Mol. Evol. 69, 164-175.
Nakashima, K., Abe, J. and Kanazawa, A. (2018) Chromosomal distribution
of soybean retrotransposon SORE-1 suggests its recent preferential insertion into euchromatic regions. Chromosome Res. 26, 199-210.
Nakashima, K., Tsuchiya, M., Fukushima, S., Abe, J. and Kanazawa, A. (2018)
Transcription of soybean retrotransposon SORE-1 is temporally upregulated in developing ovules. Planta 248, 1331-1337.
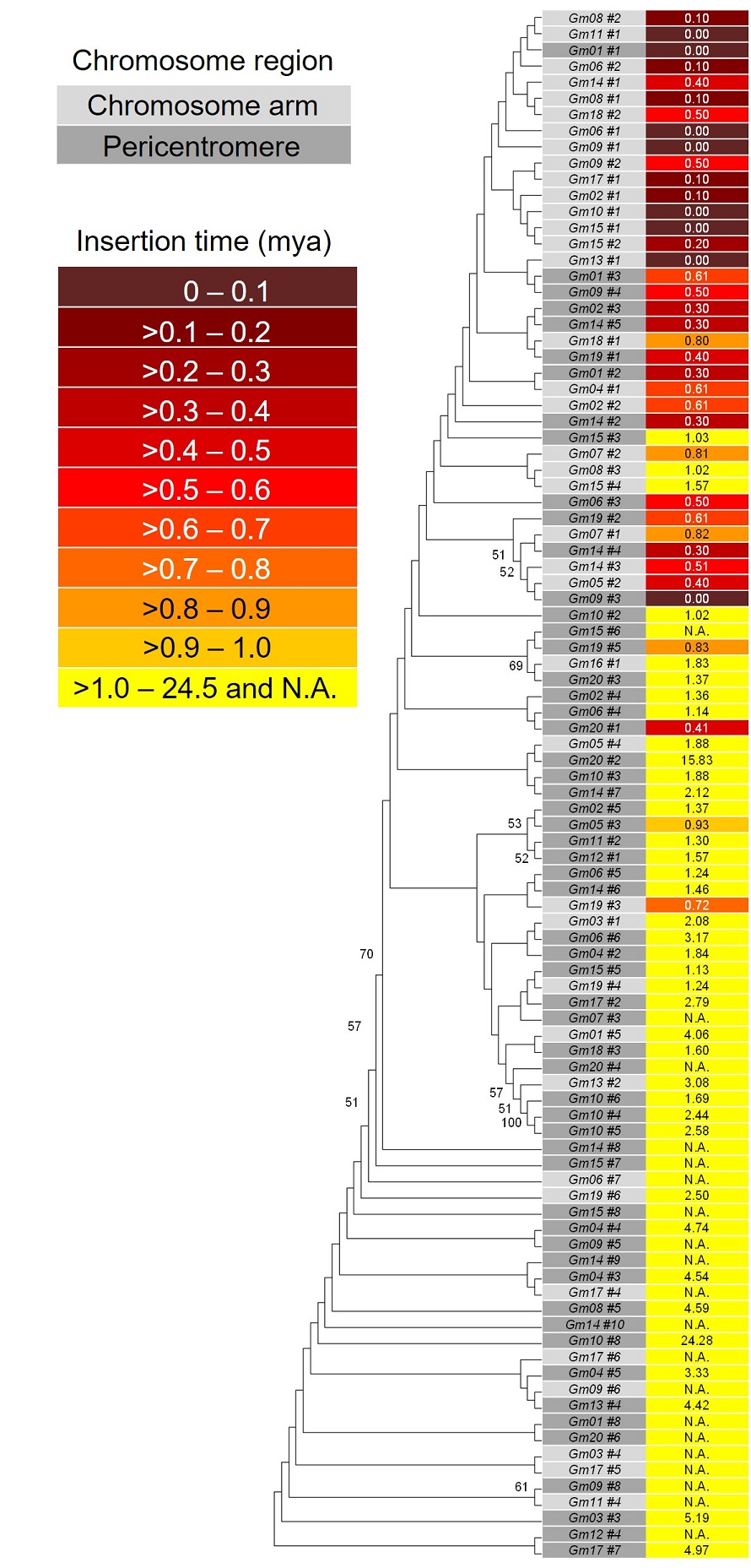
トランスポゾン(転移因子)が遺伝子内に転移して遺伝子が壊れると、多くの場合、生物の生育に不利となり、適応度が低下します。一方、数は少ないものの、転移因子が挿入することにより適応度が増大した事例が知られています。その例として、コムギにおいて開花制御遺伝子の発現増強が生じて適応性が拡大した事例や、ショウジョウバエにおいて殺虫剤に対する耐性が獲得された事例を挙げることができます。上記の論文で発表した研究では、ダイズの日長感応性の違いによって開花時期が異なる系統間の比較を行いました。その結果、ゲノム中に複数存在する、光受容体であるフィトクロームAタンパク質をコードする遺伝子の一つにレトロトランスポゾンが挿入し、その遺伝子が不活性化していることを見出しました。このことによりダイズは日長に対する感受性が低下して早く開花し、北海道のように日長が長い高緯度地方における生育に適した形質が生じていました。発見当時、このレトロトランスポゾンは新規なものであり、SOybean
RetroElement 1 (SORE-1) と名付けました。
SORE-1 が転移した先が遺伝子領域であったのは、偶然だったのでしょうか?レトロトランスポゾンが転移してからの年代に対応して塩基の変異が蓄積されることを利用して、そのレトロトランスポゾンがどれほど前に転移したものであるかを推定することが可能です。この図はゲノム中に存在するSORE-1の多数のコピーの塩基配列を基に推定した進化的な関係とそれぞれのコピーの挿入年代を表しています。解析の結果、両者の関係は関連しており、古い方のコピーの多くの挿入時期は100万年~2,450万年前(黄色)であり、新しい方は現在~10万年前(濃い赤色)でした。また、薄い灰色と濃い灰色は、それぞれ染色体上の染色体腕部とペリセントロメア領域に存在するコピーであることを示しています。染色体腕部にはペリセントロメア領域よりも相対的に遺伝子が多く存在します。したがって、この解析から、最近転移したSORE-1は遺伝子が多く存在する領域に転移していることが示唆されました。また、SORE-1の転移に必要な遺伝子発現が、次世代に伝達される遺伝情報を持つ生殖細胞を含む胚珠において増加することを見出しました。このようなことから、現在、SORE-1は遺伝的な変化を生み出す性質を持っていると考えられます。この性質を利用して、SORE-1の転移を介して有用な形質を持ったダイズの遺伝資源を作出することが可能になるかもしれません。